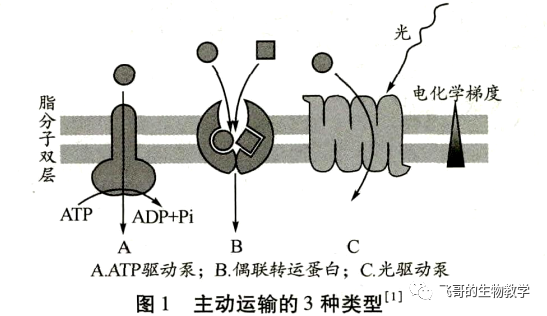
主动运输普遍存在于动植物细胞和微生物细胞中,是由载体蛋白所介导的物质逆浓度梯度或电化学梯度由低浓度一侧向高浓度一侧进行跨膜转运的方式,且转运物质的过程需要与某种释放能量的过程相偶联。一般根据能量来源,将主动运输归纳为:
由ATP 直接提供能量(ATP 驱动泵)、间接提供能量(偶联转运蛋白)和光能驱动3种基本类型(图1)。
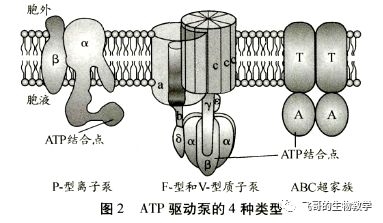
ATP 驱动泵(ATP-driven pump)是ATP 酶,直接利用水解 ATP提供的能量,实现离子或小分子逆浓度梯度或电化学梯度的跨膜运动,每秒转运的离子数为100~103不等。ATP 驱动泵都是跨膜蛋白,在膜的原生质表面具有一个或多个ATP结合位点,在逆浓度转运离子或小分子的同时水解 ATP,将储存在高能磷酸键中的能量释放出来。高中生物教学中讲的主动运输一般就是通过 ATP 水解直接或者间接供能,其中最典型的是直接水解 ATP 供能的 ATP驱动泵。根据泵蛋白的结构和功能特性,ATP驱动泵又可分为4类:P-型离子泵,V-型质子泵,F-型质子泵和ABC超家族。前3种只转运离子,后一种主要转运小分子(图2)。
Na+-K+泵是其中最常见的,也是目前研究最充分的一种ATP-驱动泵(属于P-型离子泵类型),普遍存在于动物细胞的细胞质膜上。一般动物细胞要消耗1/3 的总 ATP(神经元则要消耗2/3的总 ATP)来维持细胞内低Na+高K+的离子环境。这种特殊的离子环境对维持细胞正常的生命活动,神经冲动的传播以及维持细胞的渗透平衡,恒定细胞的体积都是非常必要的。K+和Na+逆浓度与电化学梯度输入和输出的跨膜转运是一种基本的、典型的主动运输。
例如,神经元在静息状态下,由于细胞内外K+浓度的不同和细胞膜对K+有较高的通透性(比Na+通透性高出50~100倍)促使K+外流,K+外流以建立起神经元膜外正内负的电位差阻止K+外流。当前者的力量大于后者时,K+继续外流;当二者的力量相等,则此时K+的净流动等于零,表明所建立的静息电位正好阻止K+的外流,从而使静息电位维持恒定值,K+在神经元细胞内维持在较高较稳定的浓度,此时K+浓度达到平衡。当神经元受到有效刺激时,细胞膜对Na+的通透性突然增大,Na+通过Na+通道蛋白(Na+通道蛋白由4 个亚单位环绕结合在一起形成Na+通道的孔道)迅速内流。在静息电位时,K+通过K+通道蛋白扩散到细胞外,动作电位时,Na+通过Na+通道蛋白迅速内流,此过程K+和Na+都是通过通道蛋白顺浓度梯度方向进出细胞且不耗能,属于协助扩散。形成动作电位后,神经元膜上的Na+-K+泵每消耗1分子ATP泵出3分子Na+,泵进2分子K+,通过逆浓度梯度运输维持神经元细胞内高Na+低 K+,此过程消耗能量且逆浓度梯度转运,K+泵进细胞和Na+泵出细胞都属于主动运输。
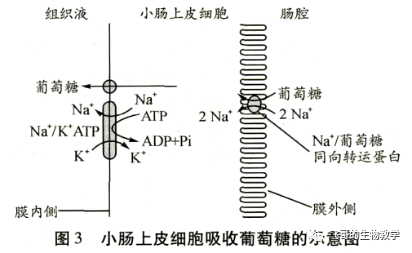
偶联转运蛋白(coupled transporter)介导各种离子和分子的跨膜运动,包括同向转运蛋白和反向转运蛋白等2种基本类型。这2类转运蛋白都使要转运的离子或分子逆浓度梯度的运动与一种或多种不同离子顺浓度梯度的运动偶联起来。偶联转运蛋白所利用的能量是由其中一种溶质的电化学梯度来提供的,而溶质的电化学梯度则依赖 ATP 的消耗来维持。因此,偶联转运蛋白介导各种离子和分子的跨膜运动是间接消耗 ATP 来实现的。例如,Na+主要是维持动物细胞外的渗透压,小肠上皮细胞和肾小管上皮细胞吸收葡萄糖,就是伴随着Na+从细胞外流入细胞内(Na+顺浓度梯度运输)完成的,而细胞膜上的 Na+-K+泵消耗 ATP 维持Na+在质膜两侧的电化学梯度,因此葡萄糖通过利用Na+的电化学梯度间接消耗ATP进入细胞(图3),此过程属于主动运输。
小肠上皮细胞的细胞膜上同向转运蛋白使葡萄糖伴随着Na+一同进入细胞内,与此同时细胞膜上Na+-K+泵通过消耗 ATP将细胞内的Na+主动转运到细胞外,当流入细胞的Na+与流出细胞的Na+的速率相等,则Na+的净流动等于零,最终维持Na+在小肠上皮细胞内的较低浓度水平。
光驱动泵(light-driven pump)主要在细菌细胞中发现,对溶质的主动运输与光能的输入相偶联,如菌紫红质利用光能驱动H+的转运。
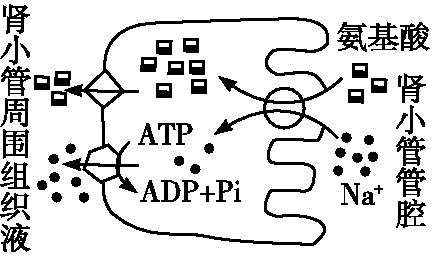
例题:如图为氨基酸和Na+进出肾小管上皮细胞的示意图。下列叙述中不正确的是( )

 网站首页
网站首页 必修一
必修一 必修二
必修二 选修一
选修一 选修二
选修二 选修三
选修三 高考竞赛
高考竞赛 教材汇总
教材汇总