最早观察染色体末端的科学家始于19世纪末期,Rabl在1885年注意到染色体上所有的末端都处于细胞核的一侧。20世纪30年代,两个著名的遗传学家McClintock B和Muller HJ 发现了染色体的末端可维持染色体的稳定性和完整性。Muller将它定义为“telomere”,这是由希腊词根“末端”(telos)及“部分”(meros)组成的。30多年前,Hayflick首次提出将体外培养的正常人成纤维细胞的“有限复制力”作为细胞衰老的表征。在此过程中,细胞群中的大部分细胞经历了一定次数的分裂后便停止了,但它们并没有死亡,仍保持着代谢活性,只是在基因表达方式上有一定的改变。于是Hayflick猜测细胞内有一个限制细胞分裂次数的“钟”,后来通过细胞核移植实验发现,这种“钟”在细胞核的染色体末端——端粒。但端粒究竟是怎样的复杂结构呢?Blackburn和Gall于1978年首次阐明了四膜虫rDNA分子的末端结构,他们发现这种rDNA每条链的末端均含有大量的重复片段,并且这些大量重复的片段多是由富含G、C的脱氧核苷酸形成的简单序列串联而成。在1985年,CWGreider和EHBlackburn发现将一段单链的末端寡聚核苷酸加至四膜虫的提取物中后,端粒的长度延长了,这就说明了确实有这样的一种酶存在,并将它命名为“端粒酶”(telomerase)。之后,耶鲁大学Morin于1989年在人宫颈癌细胞中也发现了人端粒酶。近年来,随着人体端粒酶的发现和端粒学说的提出,已经知道决定细胞衰老的“生物钟”就是染色体末端的端粒DNA,它可随着年龄的增长而缩短。
多人错误的认为,退休是一个人进入生理老年的开端。而老年则是衰老的标志,其实,这是不科学的。人体的所有器官和组织都由细胞组成,但组成器官和组织的细胞有两大类,即干细胞和非干细胞。人体衰老正是由细胞特别是干细胞衰老引起的。医学家认为,如果人类若能避免一些疾患和意外事故,人类寿命的上限应当是130岁。在人类基因组计划之前和进行之中,对长寿的分子生物学研究就有了许多显著的成果与发现。总的归纳起来便是:衰老是一种多基因的复合调控过程,表现为染色体端粒长度的改变、DNA损伤(包括单链和双链的断裂)、DNA的甲基化和细胞的氧化损害等。这些因素的综合作用,才造成了寿命的长短。
近几十年来,随着现代遗传学、分子生物学、细胞生物学和分子免疫学等边缘学科的飞速发展,人们对衰老的机理有了深层次的认识,有许多学说如遗传程序学说、DNA分子修复能力下降假说、体细胞突变学说、差错灾难学说和交联学说等已经被人们广泛接受,但端粒学说刚进入人们的研究范围。端粒缩短可引起衰老,而维持端粒长短的重要活性物质便是端粒酶。生物学家早就发现一件有趣的事实:就是每一种细胞的寿命都有一定限度,在人工培养条件下,接近这个限度时,哪怕用最好的培养方法都拯救不了既定的命运。像人体的成纤维细胞,据试验,最多只能繁殖50代,到那时必然趋于死亡。其他像老鼠的成纤维细胞只能分裂18代,龟的成纤维细胞分裂110代,如此等等。那么人为什么会衰老,以至走向死亡呢?有研究者对导致人体细胞衰老的原因提出了“程序假说”和“错误积累假说”。
人类的细胞并不能无限制地重复分裂,在分裂50~60次后便会停止。细胞不再继续分裂的机体组织,便呈现出衰老和机能低下的状态。随着细胞重复分裂使端粒缩短到一定的长度,从而使细胞停止了分裂。这就是“程序假说”。
细胞分裂的时候,DNA被复制,但是由于X射线、紫外线、活性氧、有害物质的损害,DNA会发生异常变化,于是DNA在复制过程中就会产生错误。随着错误的积累,生成了异常蛋白质,细胞机能变得低下,于是细胞便不能继续分裂,呈现出了衰老迹象。这就是所谓“错误积累假说”。
因此,人不像机器那样容易磨损和坏掉,而是能自我成长和修复,但这只能算是衰老的伴生现象。对衰老机理的研究就是为了有效地指导抗衰老的研究和实践工作。但是,人类衰老的原因是多方面的,衰老的机理也是极为复杂的。
端粒是真核细胞内染色体末端的DNA重复片断,经常被比做鞋带两端防止磨损的塑料套,由富含G的核酸重复序列和许多蛋白质组成,包括Ku70、Ku80、依赖DNA的蛋白激酶和端粒重复序列结合因子2(TRF2)等。不同个体的端粒初始长度差异很大,在人中大约为15 kb,在大鼠中可长达150 kb,在小鼠中一般在5~80 kb之间变化,而在尖毛虫中却只有20 bp。在所有的有机体中,端粒DNA的长度总是随着外界环境而波动变化的。酵母的端粒DNA在200~400 bp间随遗传或营养状态的改变而改变,四膜虫和锥虫等有机体的端粒长度在对数期会持续增加。相反,在人体中,随着细胞的持续分裂,端粒会缓慢缩短。细胞培养研究表明,当端粒再也无法保护染色体免受伤害时,细胞就会停止分裂,或者变得不稳定。其功能是完成染色体末端的复制,防止染色体免遭融合、重组和降解。染色体复制的上述特点决定了细胞分裂的次数是有限的,端粒的长度决定了细胞的寿命,故而被称为“生命的时钟”。
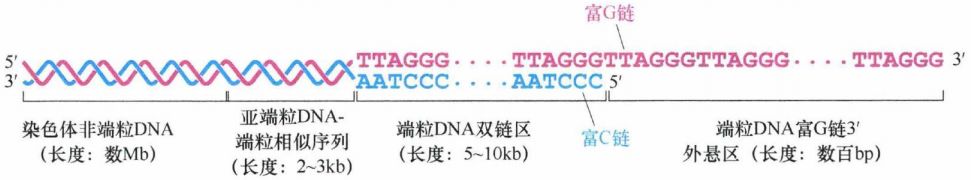
在哺乳动物细胞(以及许多其他后生动物细胞)中,端粒由重复的六核苷酸序列组成,其中一条链(富含G)上为5'-TTAGGG-3', 互补链上(富含C)为 5'-CCCTAA-3'。在正常人体细胞中,端粒DNA由数千个重复的六核苷酸序列组成,在染色体末端形成5-10kb 长的序列重复片段。
端粒DNA通常为5-10kb长。在功能性端粒DNA(中间)与非端粒染色体DNA(最左侧)之间还存在着亚端粒DNA区域。亚端粒DNA区域里含有TTAGGG类似片段,但并没有染色体末端保护功能。然而,由于亚端粒DNA含有端粒类似序列,它通常也是端粒限制性片段(TRF)的组成部分。但是只有单纯的端粒重复片段能够保护染色体DNA末端:当单纯串联重复片段的重复次数减少到12次以下时就会丧失末端保护功能。因此,即使仍然有数kb长度的TRF存在,但端粒已经丧失了阻止染色体DNA末端融合的能力。
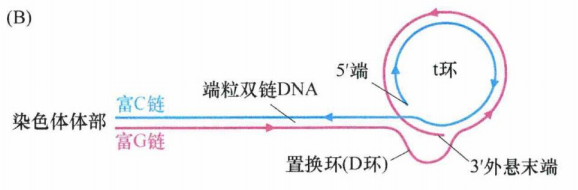
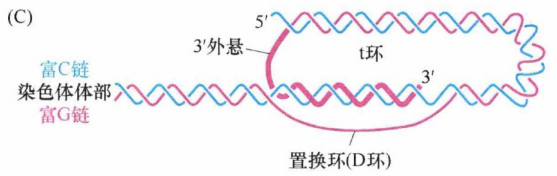
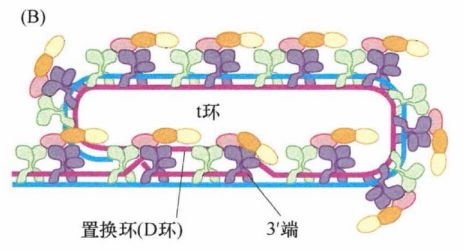
特殊的是,富G链多出一百至数百个核苷酸,导致该链3'单链端外悬。这种凸出的链会形成一种最不寻常的分子构型——t环。当时通过电子显微镜分析端粒DNA时发现了一种环形结构,实质上是套索结构。这种构型的形成依赖于三链DNA复合体的形成 。有可能所有端粒DNA的末端均含有 t 环,但是由于在电子显微镜下保存和观察此结构的技术上的限制,只有一部分端粒在电子显微镜下可以观察到 t 环。t 环有助于保护线性DNA分子未端,因为单链末端的外悬区被巧妙地塞进双链区域,以保护其免受损伤。
下图为 t 环的示意图,显示了 3'端凸出的富G链(粉色)与富C链(蓝色)的小段区域退火形成詈换(D环)(粉色链)。图中箭头方向为 5'→3'。
t 环结构
下图再次显示了t环,3'端伸入染色体形成双链螺旋,在图中用粗线标明(粉色)。
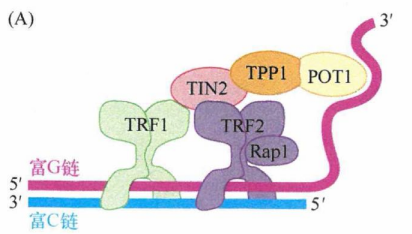
相对较长的双链端粒DNA和相对较短的外悬单链末端都与特定蛋白质相结合。这些蛋白质中含有能够特异识别并结合该六核苷酸序列的结构域,能够与端粒DNA的双链和单链区结合。端粒结合蛋白、尚未被发现的相关蛋白及端粒DNA一起构成的核蛋白复合体就称为端粒。构成端粒复合物的一些端粒相关蛋白能保护端粒免于降解。其中,TRF1和TRF2能够结合端粒的双链DNA部分,而第三个蛋白POT1既能结合富G链的 3'端外悬单链,又能与置换环(D环)的单链DNA结合。
每个端粒中会出现多个端粒复合物,它们负责t环的结构维持。如下图所示,POT1通过和D环结合能够维持其结构稳定性并维持 t 环结构完整性。
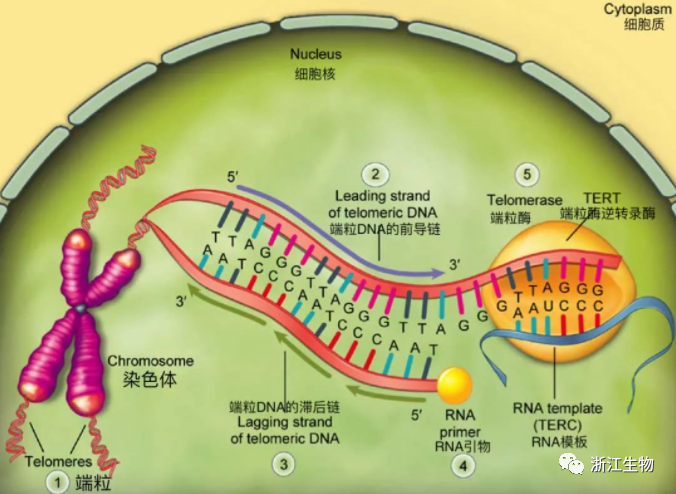
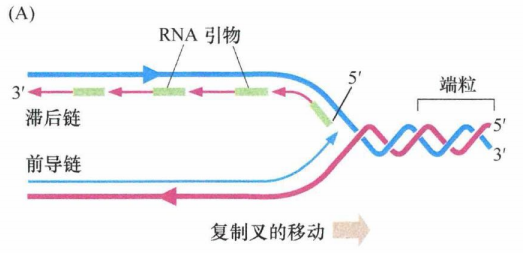
在细胞周期的S期,复制机制能高效地复制线性DNA分子中间的序列,即染色体的主体部分,但复制染色体末端序列是极其困难的。困难之处在于所有DNA链的合成必须从已有DNA的3'羟基端开始,DNA3'羟基端以与引物相似的方式来延伸核DNA链。如果没有可用的DNA引物,则以RNA分子的3'端作为DNA合成的引物。
如果引物酶(其作用是安置RNA引物)恰巧在距模板链 ( "滞后链合成”发生处)3'端的一定距离处放置了一个RNA引物,那么DNA聚合酶新合成的DNA链的 3'端将缺少一定数量与模板链互补的碱基。即使引物酶恰巧位于模板链的末端,并合成了一条RNA引物,在新合成的子链中仍缺失大约10个核苷酸,这正相当于RNA引物的长度,因为RNA在引物完成起始DNA延伸的使命后就被降解了。最后的结果是DNA两条模板链的其中一条末端的核苷酸序列不能被适当复制。
在正常DNA复制过程中,亲本的双螺旋DNA在解旋酶作用下解开,使得复制过程持续进行。新合成的滞后链由短的RNA引物片段引导,引物酶按数百个核苷酸的间隔依次安置这些RNA引物。RNA 分子的3'羟基端作为新合成子链的起始。在这些 RNA 引物尚未移去、片段尚未融合之前,这些片段称为冈崎片段。因为合成按5'→3'方向进行,所以滞后链的合成与延伸的方向和复制叉前进的方向相反。而另一个亲本链的复制由于延伸方向和复制叉的移动方向一致,能够连续进行复制,随着亲本链的解旋向3'端延伸。下图中每条链5'→3'的方向由箭头指示。粗线代表亲本DNA链,细线代表新合成链。
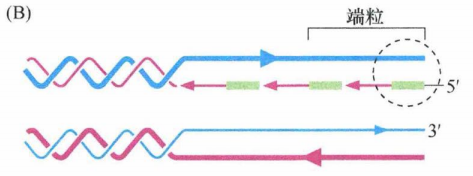
由于以上原因,前导链DNA的合成将会一直进行到端粒的末端,产生和原来端粒DNA序列一致的拷贝。然而,滞后链的合成需要用到端粒末端DNA (环状)附近对应的RNA引物。由于此RNA引物随后会被移除,而且它也未必确定位于蓝色模板链的末端精确互补,因此最终合成的DNA链是缺少一段亲本链的互补序列的,最后造成部分亲本链DNA(粗线,蓝色)没有被复制,以及子代细胞基因组中端粒DNA序列的丢失。下图中每条链5'→3'的方向由箭头指示。粗线代表亲本DNA链,细线代表新合成链。
端粒DNA的复制
末端复制的问题可以解释正常细胞每一次分裂后端粒DNA的缩短。除了端粒DNA末端的复制不足,细胞中核酸外切酶也会对端粒DNA的末端造成损坏,其最终所导致的端粒末端侵蚀也许会更大。无论何种原因,在许多类型的正常人类细胞中,细胞每分裂增殖一代端粒将会丢失50-100bp的DNA。这种对端粒DNA的持续性破坏是限制细胞传代能力的分子机制。
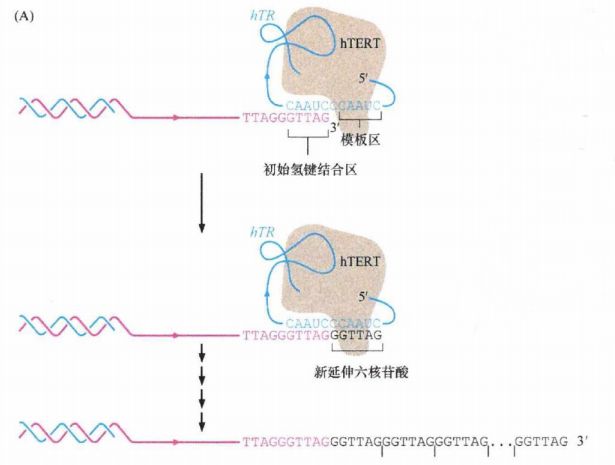
端粒酶(或端粒体酶)是一种能延长端粒末端的核糖蛋白酶,主要成分是RNA和蛋白质,其含有引物特异识别位点,能以自身RNA为模板,合成端粒DNA并加到染色体末端,使端粒延长,从而延长细胞的寿命甚至使其永生化。如果细胞被病毒感染,或者某些抑癌基因如p53、pRB等突变,细胞可越过M1期而继续分裂,端粒继续缩短,最终达到一个关键阈值,细胞进入第二致死期M2,这时染色体可能出现形态异常,某些细胞由于端粒太短而失去功能,从而导致细胞死亡。但极少数细胞能在此阶段进一步激活端粒酶,使端粒功能得以恢复,并维持染色体的稳定性,从而避免死亡。最近Shayetal在Science上发表了一幅有趣的模式图,简要介绍了端粒、端粒酶介导细胞凋亡或永生化的过程。
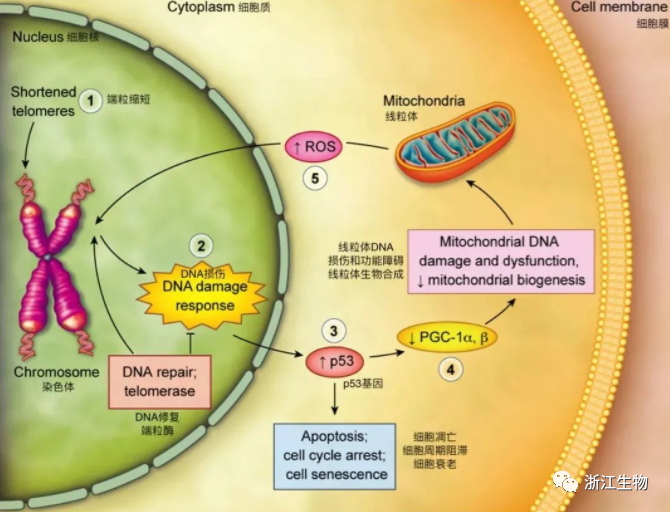
大量的证据表明,端粒酶的激活或抑制会导致细胞永生化或进入分裂终止期。端粒酶在超过80%的永生细胞系及大多数肿瘤组织中呈激活状态。端粒酶的抑制会使胚胎干细胞、骨髓造血细胞的增生受到抑制,并使肿瘤细胞系增生减弱,以致于凋亡增加。有必要指出的是:端粒酶对细胞增生、衰老及凋亡的调节是通过不同的途径进行的。其中端粒延长依赖性机制作用缓慢,需要多代细胞端粒的进行性缩短积累到一定程度,才会诱发细胞静止信号的激活。最近有一种端粒延长非依赖性机制,其作用较快,可能涉及到端粒三级结构的改变,蛋白相互作用的改变,转位的改变等。
下图中,全酶附着在3'端富含G凸出末端(粉色),部分通过hTR的氢键结合到富含G链的最后5个核酸上。因此通过对hTR亚单位序列的逆转录,hTERT能够以每次6个核苷酸的形式使富含G链延伸(黑色)。通过重复每次增加6个核苷酸这个过程,该酶能在富含G链上延伸成百上千个核苷酸。箭头方向为 5'→3', 在完成延伸后,传统的DNA聚合酶会结合到互补链(蓝色)完成DNA合成。
关于端粒丢失同衰老的关系理论是由Olovnikov博士于1973年首次提出的。他认为,端粒的丢失很可能是因为某种与端粒相关的基因发生了致死性的缺失。目前认为,人类细胞内端粒酶活性的缺失将导致端粒缩短,每次丢失50~200个碱基,这种缩短使得端粒最终不能被细胞识别。端粒一旦短于“关键长度”,就很有可能导致染色体双链的断裂,并激活细胞自身的检验系统,从而使细胞进入M1期死亡状态。随着端粒的进一步丢失,将会发生染色体重排和非整倍体染色体的形成等错误,这将导致进一步的危机产生,即M2期死亡状态。当几千个碱基的端粒DNA丢失后,细胞就停止分裂而引起衰老。端粒及端粒酶涉及衰老最有力的证据是Bodnar等证实的。如果细胞试图要维持其正常分裂,那么就必须阻止端粒的进一步丢失,并且激活端粒酶。Cooke等认为,由于人体细胞中的端粒酶未被活化,从而导致了端粒DNA缩短。因此,只有那些重新获得端粒酶活性的细胞才能继续生存下去,对于那些无法激活端粒酶的细胞将只能面临趋向衰老的结果。研究人员最近还发现,患有一种可加速衰老的遗传病人具有异常短的端粒,这进一步表明端粒在衰老过程中所起的重要作用。在人类细胞中,研究者还发现,端粒缩短的速率与细胞抗氧化损伤的能力相关。更容易遭受氧化损害的细胞,其端粒缩短更快,然而那些更能抵抗这种损伤的细胞,端粒缩短得较慢。如果能减免细胞损伤或激活端粒酶,即可控制人类的衰老进程。
有人曾经对人淋巴细胞的衰老性变化与其端粒长度以及端粒酶活性的关系在各种体内体外环境及处理因素下做了观测,发现端粒酶活性和端粒长度的调节有可能是淋巴细胞增殖的控制因素,这已在人体淋巴细胞的发育、分化、激活和衰老过程中被验证。曾发现外周血CD+4T细胞的端粒长度在体内随着衰老以及从静息细胞到记忆细胞的分化过程而缩短,在体外则随着细胞的分裂而缩短,这些结果提示端粒长度与淋巴细胞增殖过程以及记忆性增殖潜力相关。
端粒酶的表达已知能够抑制衰老,而Weinberg and colleagues认为端粒酶的作用主要在于延长了端粒悬垂的长度。细胞的复制期限被认为由最终导致衰老的两个机制决定,一个是累积的DNA损伤,另外一个是端粒的进行性缩短。Weinberg and colleagues研究了一个端粒的特殊悬垂结构在衰老过程中的作用,悬垂结构只在富含C的末端之外还有一个由几百个核苷酸组成的富含G的结构。据称Shay实验小组的研究策略是通过抑制端粒酶活性,从而迫使永生化细胞转变为正常细胞,进入正常的衰老和死亡模式。
在衰老异常发展中有一种早衰人群,即从20岁开始皮肤和毛发等便迅速衰老,其原因仍在于制造端粒酶的遗传基因。细胞在分裂的时候,DNA双螺旋结构以其一根长链为“模子”进行DNA复制。在DNA修复损伤的时候,“拆解”DNA的双螺旋结构是必要的,制造端粒酶的遗传基因在解开DNA螺旋结构上起作用。像制造端粒酶并从事DNA复制和修改错误的一类遗传基因,若与延长细胞寿命的端粒酶良好结合,我们也许能期待向“长生不老”的目标进一步接近。
总之,人类体细胞在复制衰老过程中产生的端粒丢失现象已在体外得到了证实,而且体内的端粒丢失可作为判断供体年龄的依据。我们只要设法使已衰老的人体内各种干细胞的端粒长度恢复到年轻时的水平,老人就会返老还童和长生不老。但在人类端粒及端粒酶的基础研究中,还存在着许多难点,如:人端粒末端的精细结构,端粒的非端粒酶延伸机制;人端粒酶的具体结构及其基因所在的位置;端粒酶的激活机制及其活性调节等,均有待于回答。尽管如此,我们似乎仍看到了前景的美好。毕竟人们已找到了同衰老有着紧密相关性的因素——端粒和端粒酶。人们对于端粒抑制剂的研究已经蓬勃的展开了。故进一步研究端粒酶的活性调节机制,对于开发新型延缓衰老的端粒酶抑制剂无疑具有重要意义。Colorado大学的两位研究人员Thomas Cech 和Robert Weinbrg博士已独立地克隆出一种控制人类细胞端粒酶活性的基因。应用这种基因,很有可能得到一种新的蛋白质——端粒酶的控制剂。
关于衰老机理和抗衰老的研究领域现在仍然是非常活跃的,并将受到越来越足够的重视,因为它对于延缓衰老,实验老年医学研究的目的即防止人类早衰,保持人体健康长寿是极为重要的。但是,就目前人类在这方面的研究来看还很薄弱。在今后一个时期内,有关衰老与抗衰老的研究重点还应放在以最新生物学技术研究有关长寿与衰老基因的克隆、结构分析以及对这些基因的调控机制;机体衰老过程中自由基、突变以及其它有害刺激因素启动细胞衰老凋亡的分子机制和这些过程被调控的分子机理;利用衰老基因与长寿基因的研究成果进行的基因治疗方面研究等。
1.端粒是染色体两端一段特殊序列的DNA-蛋白质复合体,端粒DNA序列随细胞分裂次数增加而缩短,当短到一定程度时,端粒内侧的正常基因会受到损伤,导致细胞衰老。端粒酶以其携带的RNA为模板(含短重复序列5'-UAACCC-3')使端粒DNA序列延伸,作用机理如图所示。下列叙述正确的是( )

A.图示端粒酶向右移动完成C链的延伸
B.端粒酶中RNA上相邻的3个碱基组成一个密码子
C.端粒酶延伸端粒DNA的短重复序列为5'-GGGTTA-3'
D.端粒酶的检测及激活可用于肿瘤的诊断和治疗
解析:A项是端粒酶向右移动完成G链的延伸,再利用G链合成C链。可根据DNA子链延伸方向只能从5'→3'来判断先合成G链。
B项端粒酶中RNA并非mRNA,不具有密码子功能。
C项根据碱基互补配对原则即可得出此项正确。
D项端粒酶激活后细胞会不断分裂难以衰老,不利于癌细胞死亡和治疗。
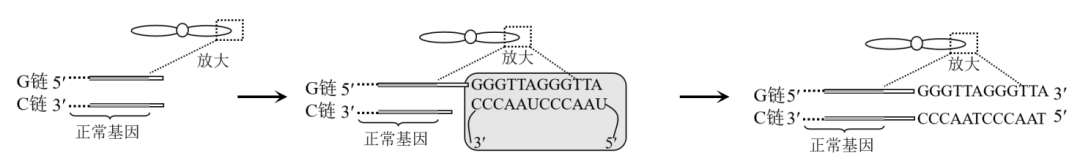
答案:C
2.人体细胞中的染色体DNA会随着复制次数增加而逐渐缩短。在生殖系细胞和癌细胞中存在端粒酶(由RNA和蛋白质形成的复合体),能够将变短的DNA末端重新加长。端粒酶作用机理如图所示。下列相关叙述错误的是( )
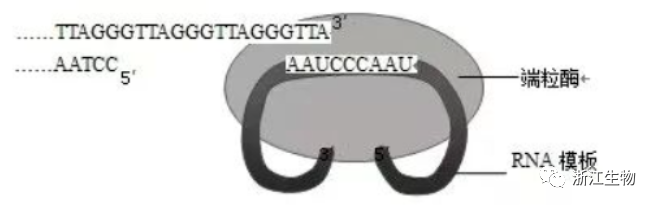
A.人体生殖系细胞以外的其他细胞不含端粒酶基因
B.端粒酶中的蛋白质能够催化染色体DNA的合成
C.细胞衰老与染色体DNA随复制次数增加而缩短有关
D.抑制端粒酶的作用可抑制癌细胞增殖
解析:人体生殖系细胞以外的其他细胞也含有端粒酶基因,没有端粒酶活性;端粒酶能以自身的RNA为模板合成端粒DNA的一条链,说明端粒酶中蛋白质为逆转录酶,能够催化染色体DNA的合成;根据细胞衰老的端粒学说,细胞衰老与染色体DNA随复制次数增加而缩短有关;端粒酶能够将变短的DNA末端重新加长,使得癌细胞能无限增殖,抑制端粒酶的作用可抑制癌细胞增殖。
答案:A

 网站首页
网站首页 必修一
必修一 必修二
必修二 选修一
选修一 选修二
选修二 选修三
选修三 高考竞赛
高考竞赛 教材汇总
教材汇总