细胞增殖的内容是《普通高中生物学课程标准(2017年版)》中的一个重要概念,也是高中生物学教材中必修模块的重点内容。有丝分裂是真核生物细胞主要的增殖方式,而减数分裂又是特殊方式的有丝分裂,所以对有丝分裂相关知识的明晰尤为重要,有助于提升学生生命观念和科学探究素养。有丝分裂最重要的变化是,在纺锤体作用下将亲代细胞复制的染色体平均分配到两个子细胞中,从而保持了细胞在遗传上的稳定性。但在教学中,部分教师对细胞分裂中纺锤体的形成以及与其有关的秋水仙素,存在困惑。鉴于此,笔者从教学实践出发,围绕“纺锤体”这一名词,对有丝分裂中几个重要知识点进行阐释,希望将生物学的科学本质传递给学生,更好地发展学生的科学思维。
1纺锤体是如何形成的
纺锤体是动植物细胞分裂期形成的与染色体正常分离直接相关的分裂器,纺锤体的装配在有丝分裂的前期完成。动物细胞纺锤体由星体微管、极间微管、动粒微管及其结合蛋白构成,因含有星体微管故称有星纺锤体。无中心体的动物细胞和植物细胞也能形成纺锤体,因不含有星体微管而称之为无星纺锤体。微管是由α、β微管蛋白异源二聚体及少量微管结合蛋白聚合而成的亚稳定动态结构。
动物细胞的中心体由一对相互垂直的圆筒状中心粒及中心体基质构成。它是纺锤体微管向外生长的核心,又称微管组织中心。在有丝分裂前间期的S期初期,中心体开始复制倍增,在G2期结束时完成。在细胞分裂期前期,间期复制倍增的两个中心体分离,每一个中心体形成放射状排列的微管,称为星体,每个中心体是它自身星体的核心。
在有丝分裂细胞周期的分裂期,微管通过持续增加和丢失组成微管的微管蛋白亚基来实现微管的聚合和解聚,微管始终处于生长和缩短的更替中。在分裂前期,纺锤体微管由游离的微管蛋白组装而成,介导染色体的运动;分裂末期,纺锤体微管解聚,又组装形成细胞质微管网络。纺锤体微管包括动粒微管、极间微管和星体微管(图1)。
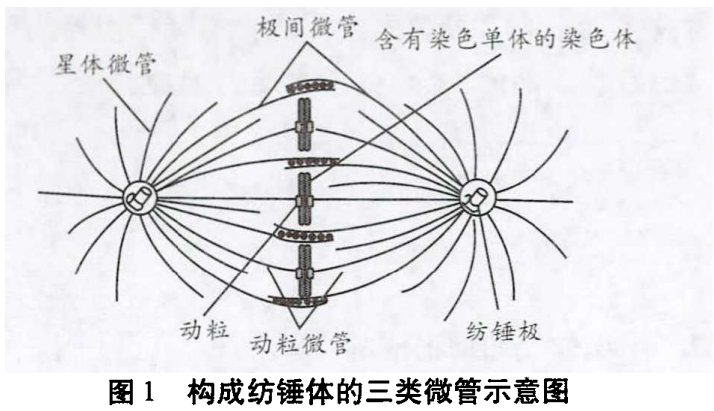
动粒微管连接染色体动粒与位于两极的中心体。在有丝分裂前期,一旦核被膜解聚,由相反两个方向的中心体伸出的动粒微管就会随机地与染色体上的动粒结合而俘获染色体,微管最终附着在动粒上,动粒微管把染色体和纺锤体连接在一起。在细胞分裂期的后期,分开后的染色单体被拉向两极。染色体移动由两个相互独立且同步进行的过程所介导,分别为过程A和过程B。在过程A中,在连接微管和动粒的马达蛋白的作用下,动粒微管解聚缩短,在动粒处产生的拉力使染色体移向两极。极间微管是从一个中心体伸出的某些微管与从另一个中心体伸出的微管相互作用,阻止了它们的解聚,从而使微管结构相对稳定,两套微管的这种结合形成了有丝分裂纺锤体的基本框架,具有典型的两极形态,产生这些微管的两个中心体称为纺锤极,这些相互作用的微管被称为极间微管。在有丝分裂后期过程B中,极间微管的伸长和相互间的滑行使纺锤极向两极方向移动。星体微管从中心体向周围呈辐射状分布,在有丝分裂后期过程B中,每一纺锤极上向外伸展的星体微管发出向外的力,拉动两个纺锤极向两极方向移动。
综上,不管是动物细胞还是植物细胞,在有丝分裂的前期都会形成纺锤体,构成纺锤体的微管化学本质都是微管蛋白,纺锤体微管在染色体的移动和分离中起着重要作用。动粒微管附着于染色体上的动粒以及纺锤体的装配还是细胞周期调控中的重要检查点。
2构成纺锤体的是纺锤丝还是星射线
人教版《生物·必修1·分子与细胞》第6章在讲述有丝分裂时,关于动物细胞和植物细胞纺锤体形成的区别是这样描述的:植物细胞是从细胞的两极发出纺锤丝,形成一个梭形的纺锤体。而动物细胞是在两极的中心粒周围发出大量的星射线,两组中心粒之间的星射线形成了纺锤体。而在《生物·必修2·遗传与进化》第2章以哺乳动物精子形成过程为例讲述减数分裂过程时,又用了“纺锤丝”这一表述。同一套教材,前后表述不一致,让教师的教学和学生的学习都产生了困惑。“纺锤丝”一词的由来是因为纺锤体微管在电子显微镜下呈丝状,在浙科版教材中即为这样表述,且不论动物细胞还是植物细胞都用“纺锤丝”。不管是纺锤丝还是星射线,都是教材编写者为了学生更好地理解和学习“纺锤体微管”这一名词。但笔者认为,要培养学生的科学思维,就应该培养学生用专业名词来描述生物学现象的能力,也能做到中学教材与大学教材内容的一致性。对于学生来说,纺锤体微管、纺锤丝和星射线都属千陌生名词,建议在教材修订时统一用“纺锤体微管”这一表述,如果担心学生对“微管”这一名词不太容易接受,可以参照浙科版教材,在细胞骨架这一部分内容增加对微管和微丝的简单介绍。即使教材仍然沿用纺锤丝或星射线这一概念,建议中学教师在教学中也能适当地给学生讲述“纺锤体微管”这一本质概念,培养学生的科学思维。
3秋水仙素会使动物细胞染色体加倍吗
微管蛋白按照来源可分为植物微管蛋白和动物脑蛋白。因植物微管蛋白难以制备,秋水仙碱与动物脑微管蛋白结合反应研究得要更多一些。秋水仙碱是从植物秋水仙中提纯出的一种生物碱,又名秋水仙素,构成微管的α、β微管蛋白异源二聚体是秋水仙素分子的结合靶点。当秋水仙碱与正在进行有丝分裂的细胞接触时,秋水仙碱结合到
微管蛋白的特定位点,导致α微管蛋白与β微管蛋白二聚体结构变形,从而阻断微管蛋白组装成微管,但并不影响微管蛋白的解聚,所以纺锤体会迅速消失。
秋水仙碱的浓度和作用时间对动、植物细胞染色体加倍的影响是关键。赵鑫闻等对美洲黑杨花粉母细胞染色体加倍的有效处理时期及有效处理浓度进行了研究。结果表明,在花粉母细胞减数分裂细线期与粗线期进行美洲黑杨2n花粉的诱导效果最好,总体上在减数分裂粗线期进行诱导得到的2n花粉最多,并且诱导的最佳浓度为0.5%。刘爱生等在利用人类外周血淋巴细胞进行染色体G显带制作中,在阻断培养的4h内任意时间加入相应剂量的秋水仙素,能获得用于G显带的形态完好、大小适中、分散均匀、轮廓清楚的中期染色体标本相。陈长超等利用秋水仙碱处理MⅠ期卵母细胞,结果发现Ml期纺锤体发生解聚,染色体周围纺锤体微管全部消失或部分残留,染色体排列异常。
综上,用适宜浓度的秋水仙素在适宜的时间作用于动物细胞和植物细胞,都会与纺锤体微管特异位点结合从而阻断纺锤体的组装,并能够引起已经组装的纺锤体解聚消失,从而使正在分裂的动、植物细胞染色体数目加倍或其他异常,并阻滞细胞分裂周期的正常进行。
4秋水仙素为什么会使有丝分裂的细胞停滞于中期
如果用秋水仙素处理有丝分裂的细胞,纺锤体会迅速消失,细胞停滞在有丝分裂中期,染色体无法分离成两组。用秋水仙碱进行诱导,从而将细胞阻断在细胞分裂中期,也是诱导细胞周期同步化的重要方法之一。很多教师和学生在教学中都会有困惑,纺锤体的组装是在细胞分裂的前期,为什么用秋水仙素处理细胞分裂不是停滞在前期而是中期?
笔者认为,这与细胞周期运转调控密切相关。真核细胞周期可分为4个时期,分别是G1期、S期、G2期和M期。在细胞周期调控中主要有3个控制点,第一个控制点在G1期,决定细胞能否进入S期;第二个控制点在G2期,决定细胞能否进入有丝分裂期;第三个控制点在M期,决定细胞是否已经准备好将复制好的染色体拉向两极。
CDK(周期蛋白依赖性蛋白激酶)对细胞周期运行起着核心性调控作用,CDK与不同时期的周期蛋白结合会在特定周期起调节作用。例如,与M期周期蛋白结合的CDK在M期起调节作用。cyclinA、cyclinB是在M期起调节功能的两种主要周期蛋白。细胞周期运转到分裂中期后,在后期促进复合物(APC)的作用下,M期cyclinA和cyclinB通过泛素化途径迅速降解,Cdkl活性丧失,细胞周期便从M期中期向后期转化。APC活性变化是细胞周期由分裂中期向后期转换的关键因素,其活性受到多种因素的综合调节,纺锤体组装检查点是其重要的调控因素。纺锤体组装不完全,或所有动粒不能被动粒微管全部捕捉,则APC不能被激活。
综上,用秋水仙素处理有丝分裂的细胞,纺锤体微管因不能组装而无法捕捉到动粒,APC无法被激活,导致cyclinA和cyclinB不能降解,Cdkl仍然保持活性,靶蛋白无法去磷酸化,M期中期无法向后期转化,从而使细胞停滞在中期。但有资料表明,去除秋水仙素的影响,细胞又会恢复纺锤体的组装,从而使细胞完成细胞周期。笔者认为,细胞停滞在中期应该是从分裂时相的角度来说的,也是染色体螺旋化程度最高的时期。作为中期的标志性特征之一,正常情况下所有染色体都整列在细胞中央的赤道板上,用秋水仙素处理后,将会很难整齐排列在赤道板,在细胞中会散乱分布。
参考文献:
[1]杨忠顺.“细胞分裂”教学中的“纺锤体”释疑[J].中学生物教学,2021(28):45-47.

 网站首页
网站首页 必修一
必修一 必修二
必修二 选修一
选修一 选修二
选修二 选修三
选修三 高考竞赛
高考竞赛 教材汇总
教材汇总